
梅斯医学MedSci APP
医路相伴,成就大医



梅斯医学MedSci APP
医路相伴,成就大医
长链非编码RNA(lncRNA)是一类转录本长度超过200nt的RNA分子,它们并不编码蛋白,而是以RNA的形式在多种层面上(表观遗传调控、转录调控以及转录后调控等)调控基因的表达水平。
lncRNA 起初被认为是基因组转录的“噪音”,是RNA聚合酶II转录的副产物,不具有生物学功能。然而,近年来的研究表明,lncRNA参与了X染色体沉默,基因 组印记以及染色质修饰,转录激活,转录干扰,核内运输等多种重要的调控过程,lncRNA的这些调控作用也开始引起人们广泛的关注。哺乳动物基因组序列中 4%~9%的序列产生的转录本是lncRNA(相应的蛋白编码RNA的比例是1%),虽然近年来关于lncRNA的研究进展迅猛,但是绝大部分的 lncRNA的功能仍然是不清楚的。
生物学功能
许多lncRNA都具有保守的二级结构,剪切形式以及亚细胞定位,这种保守性和特 异性表明它们是具有功能的。但lncRNA的功能相对于microRNA和蛋白质的功能来说更加难以确定,因为目前并不能仅根据序列或者结构来推测它们的 功能。根据它们在基因组上相对于蛋白编码基因的位置,可以将其分为(1) sense, (2) antisense, (3) bidirectional, (4) intronic, (5) intergenic这5种类型。这种位置关系对于推测lncRNA的功能有很大帮助。
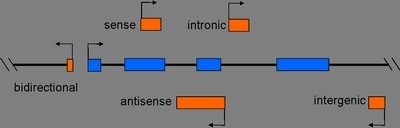
图1. 根据lncRNA在基因组上的位置,可将其分为5种类型:1. sense, 2. antisense, 3. bidirectional, 4. intronic, 5. intergenic。图中编码RNA和非编码RNA外显子分本别用蓝色和红色表示。
近年来通过对已发现的lncRNA的研究表明,lncRNA能够在多种层面调控基因的表达水平,其调控机制开始为人们所揭示(图2)。

图2. lncRNA的作用机制。根据今年来所发现的lncRNA的作用机制,lncRNA主要可能具有以下几个方面的功能:1)通过在蛋白编码基因上游启动子区 (桔)发生转录,干扰下游基因(蓝)的表达(如酵母中的SER3基因)。2)通过抑制RNA聚合酶II或者介导染色质重构以及组蛋白修饰,影响下游基因 (蓝)表达(如小鼠中的p15AS)。3)通过与蛋白编码基因的转录本形成互补双链(紫),进而干扰mRNA的剪切,从而产生不同的剪切形式。4)通过与 蛋白编码基因的转录本形成互补双链(紫),进一步在Dicer酶作用下产生内源性的siRNA,调控基因的表达水平。5)通过结合到特定蛋白质 上,lncRNA转录本(绿)能够调节相应蛋白的活性。6)作为结构组分与蛋白质形成核酸蛋白质复合体。7)通过结合到特定蛋白上,改变该蛋白的胞质定 位。8)作为小分子RNA,如miRNA,piRNA的前体分子转录(Jeremy E. Wilusz et al, 2009, Genes Dev.)。
一般来说,lncRNA主要从以下三种层面实现对基因表达的调控:
1. 表观遗传学调控
lncRNA 招募染色质重构复合体到特定位点进而介导相关基因的表达沉默。例如来源于HOXC基因座的lncRNA HOTAIR,它能够招募染色质重构复合体PRC2并将其定位到HOXD位点,进而诱导HOXD位点的表观遗传学沉默。同 样,Xist,Air,Kcnq1ot1这些lncRNA都能够通过招募相应的重构复合体,利用其中的甲基转移酶如Ezh2或者G9a等实现表观遗传学沉 默。
2. 转录调控
lncRNA能够通过多种机制在转录水平实现对基因表达的沉默, 表现在如下几个方面:lncRNA的转录能够干扰临近基因的表达。例如在酵母中,SER3基因会受到其上游lncRNA SRG1的转录的干扰;lncRNA能够通过封阻启动子区域来干扰基因的表达。例如,DHFR上游的一个lncRNA能够和DHFR的启动子区域形成 RNA-DNA3螺旋结构,进而抑制转录因子TFIID的结合,从而抑制DHFR的基因表达;lncRNA能够与RNA结合蛋白作用,并将其定位到基因启 动子区从而调控基因的表达。例如,CCND1启动子上游一个lncRNA能够调节RNA结合蛋白TLS的活性,进而调控CCND1的表达;lncRNA能 够调节转录因子的活性,里例如lncRNA Evf2能够与转录因子Dlx2形成转录复合体从而激活Dlx6的表达;lncRNA也能够通过调节基本转录因子来实现调控基因的表达。例如,Alu RNA能够通过抑制RNA聚合酶II来实现广谱的基因抑制。
3. 转录后调控
lncRNA 能够在转录后水平通过与mNRA形成双链的形式调控基因的表达。例如,Zeb2 antisense RNA能够和Zeb2 mRNA内含子5’剪切位点区域形成双链,从而抑制该内含子的剪切。而该区域含有对于Zeb2蛋白表达所必须的核糖体结合位点,Zeb2 antisense RNA通过这种方式,能够提高Zeb2蛋白的表达量。
LncRNA与疾病
大量的研究表明,在肿瘤细胞中,某些特定的lncRNA的表达水平会发生改变。这种表达水平的变化能够作为癌症诊断的标志物(有时是非常灵敏的诊断标志物,如前列腺癌中的DD3,表1)和潜在的药物靶点(图3)。

图3. 近来在对阿兹海默症的研究中找到的一个lncRNA,BACE1AS,它编码β分泌酶基因的反义链RNA。β分泌酶能够产生β淀粉样蛋白,后者的累积是阿 兹海默症的主要诱因。作为BACE1反义链的BACE1AS能够在各种外界压力刺激条件下,增加BACE1 mRNA的稳定性(通过防止BACE1受到核酸酶降解的方式),从而导致更多的β淀粉样蛋白累积,并促进BACE1AS的表达,这个正反馈循环将会加速阿 兹海默症的发展。但是,当使用了特异性针对BACE1AS的siRNA降低BACE1AS的表达水平后,β淀粉样蛋白的表达水平也同时下降了,这表明 BACE1AS是一个非常理想的治疗阿兹海默症的药物靶点(Mohammad Ali Faghihi, et al. 2008. Nature Medicine)。
{nextpage}



2012年2月12日,Nature发表了一篇题为“Combined RNAi and localization for functionally dissecting long noncoding RNAs”的文章,Frank Buchholz等研究人员利用c-KLAN技术对长链非编码RNA(lncRNAs)进行了功能鉴定和定位,该技术为检测lncRNAs提供了一种高效、可靠的检测方法。
尽管当前有多种方法可用于全面研究蛋白质编码基因的细胞内作用,然而用于系统研究与多种生物学信号通路有关的长链非编码rnas (lncRNAs)的技术却十分有限。在这篇文章中,来自德国德累斯顿工业大学的研究人员开发了一种结合非编码RNAs敲除和定位分析的技术,称之为c-KLAN。利用这一技术对lncRNAs进行了功能鉴定和定位,并鉴别出了调控小鼠胚胎干细胞类型的转录物。
在真核生物基因组中大量的蛋白质编码和非编码转录物被转录,然而直到近年来后者才获得广泛的科学关注。LncRNAs是一类非蛋白质编码转录物,长度在几百bp到几kb之间。 众所周知其参与了多种生物学过程,例如X-染色体失活、染色质水平的表观遗传学状态调控、胚胎干细胞状 态维持、转录调控以及疾病状态调控等。尽管利用标记和成像技术结合功能缺失研究已成功地应用于大量的蛋白质编码基因研究中,却还没有关于长链非编码转录物 显像和表型特征鉴定的类似方法被报道。核酸内切酶切割制备的短干扰RNAs (endoribonucleaseprepared short interfering RNAs,esiRNAs)已被证实更加适合于RNAi筛选,因为它们能够有效地敲除靶向转录物,又避免了天然复杂的siRNAs的显著脱靶效应。研究人 员认为相同的优势同样适用于沉默lncRNAs。
在这篇文章中,研究人员构建了一个能靶向594种lncRNAs的esirna文库,在小鼠细胞中进行了功能缺失筛查。同时,研究人员还改良了esiRNA合成方法,实现了无缝、可再生合成特异性探针检测了细胞中lncRNAs的定位。

Incrnas的esiRNAs和RNA FISH探针的合成路线
步骤:对lncRNA序列进行DEQOR优化和经由电脑模拟设计引物,随后对cDNA进行PCR扩增,完成对PCR扩增产物的测序和定量检测。接下来一方 面通过体外转录PCR扩增产物生成dsRNA,然后利用Rnse III酶切就生成了可用于IncRNAs功能缺失研究的esiRNA文库。另一方面可利用链特异性rna聚合酶和荧光标记UTPs对PCR进行体外转录,即可生成正义或反义探针。然后采用FISH技术对IncRNAs进行定位分析。
利用c-KLAN研究了Panct1在小鼠ESCs中的作用
研究人员利用c-KLAN揭示了小鼠胚胎干细胞中转录物Panct1的定位和功能。证实Panct1主要定位于细胞核中,其在ESC状态维持中发挥着关键性的作用。
lncRNA生物学的快速发展需要对这些转录物进行深入的检测,但由于转录组的复杂性和适当注解的局限性很难对这些转录物进行标记。在此项研究中,科研人 员介绍了一种组合方法可大规模地对lncRNAs进行定位和功能分析。c-KLAN为我们提供了一种快速、简便和可靠的途径检测lncRNA介导的各种细 胞过程的调控。
长链非编码RNA主要作用是调控基因的表达水平,并不表达蛋白,近年来的研究表明长链非编码RNA也参与了肿瘤的调控。
{nextpage}
为什么长链非编码RNA过去被忽视了呢?展望
相对于蛋白编码序列以及小分子RNA,lncRNA的研究还仅仅只是处于起步阶段,其功 能与调控机制仍有待进一步阐明。目前研究成果所展现出的lncRNA繁多的分子生物学功能,如调节转录模式,调控蛋白活性,改变RNA的剪切模式等等,为 人们提出了一个从未涉足的调控领域。
当下lncRNA的主要研究方向仍然是通过原位杂交技术,过表达技术,siRNA介导的基因沉默技术来 发现更多新的lncRNA,为目前的调控模式提供更多的支持和完善。这种传统的手段固然精确,然而却缺乏效率,随着更多高通量筛查技术的发展,如 Microarray芯片杂交技术,新一代高通量测序技术,结合生物信息学的预测工具,人们将能够更快更有效率的发现那些具有重要调控功能的 lncRNA。
